The Mormyrid Weakly Electric Fish: Gnathonemus petersii
The two major groups of weakly electric fish, the South American order Gymnotiformes and the African order Osteoglossiformes, independently developed electrosensory systems through convergent evolution (Heiligenberg, 1990). Both orders generate electric fields from electric organs that function in a manner similar to conventional batteries. This electric organ discharge is used both for communication with conspecifics and active electrolocation of objects in the fish’s environment. Electrical signals caused by external current sources, termed ex-afferent signals, are processed by either the Knollenorgan pathway, in the case of high frequency EOD emissions by conspecifics, or the ampullary pathway, in the case of low frequency signals not coincident with the fish’s own EOD. Reafferent sensory signals, those caused by distortions of the fish’s own EOD, are transduced by mormyromast receptors and processed by a third, initially separate pathway.
The neural substrate for active electrolocation, the detection of reafferent signals by the mormyromast modality, is of particular interest both because it is the most poorly understood of the three electrosensory modalities and because it involves multiple neural codes and interactions with a corollary discharge copy of the EOD motor command at the first post-receptor synapse. The active electrosensory system is an excellent example of low-level active sensory processes for these reasons (Bell, 1989).
In the active electrosensory system of the family Mormyridae, there are two separate mormyromast electroreceptor subtypes on the skin of the fish that project to different areas of the ELL, an accessory cerebellar structure devoted to the processing of electrosensory information from all three modalities (Bell, 1989). Projections from the A-type mormyromast receptors, which code intensity of electrical current inward across the skin, are sent to the medial zone of ELL. There is a precise relationship between current intensity and the timing of the afferent response of the receptor. High current intensities result in a nearly immediate response, whereas lower current intensities produce a graded delay in the generation of an action potential in the afferent. Thus, distortions in the fish’s own electric field, caused by environmental objects with certain impedances, are represented as a latency code in the primary afferent projection to the ELL (Bell, 1989).
A latency code makes the most functional sense when the cell in question delivers only a single spike (i.e., action potential) within the critical temporal window for the post-synaptic cell. A consistent, monotonically decreasing relationship between stimulus amplitude and delay to spike supports the idea of a latency code (Szabo and Hagiwara, 1967).
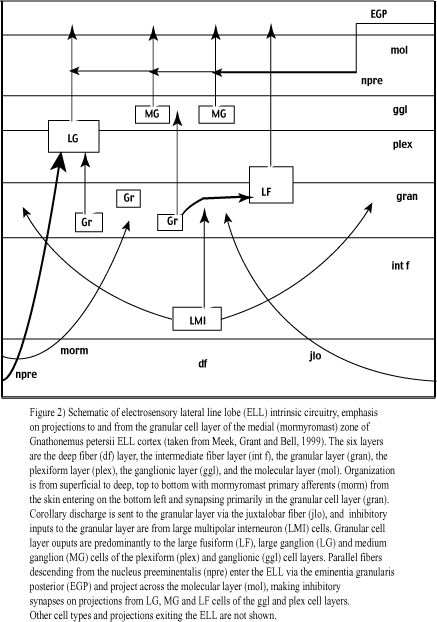
The ELL consists of six cell layers. In order from superficial to deep, these are the molecular, ganglionic, plexiform, granular, intermediate and deep fiber layers (Meek, Grant and Bell, 1999; figure 2). Inputs to the ELL include electroreceptor afferents, electric organ corollary discharge (EOCD) from the juxtalobar nucleus, proprioceptive information (for instance, tail bending influences the shape of the emitted electric field),
and feedback from higher brain areas. Outputs are primarily to the midbrain torus semicircularis (figure 2). Fourteen cell types, including GABAergic interneurons (large multipolar interneurons, LMI cells), large fusiform and ganglion projection cells, and compact granular cells with uncharacterized projections, have been identified in the medial mormyromast zone of the ELL in Gnathonemus petersii (Meek, Grant and Bell, 1999; see figure 2).
Understanding of neuronal physiology and active signaling allows us to meaningfully inquire into the function of ELL cells and their connections. Neurons summate voltage inputs from other neurons in a graded manner to produce a digital, all-or-none depolarizing event, termed an action potential or spike, if a threshold level of depolarization is reached. The spike is then propagated down long axonal fibers to presynaptic terminals. At these terminals, or boutons, the all-or-none spike is converted to a graded-intensity signal at the postsynaptic neuron following either chemical or electrical transmission. Corresponding postsynaptic events are either excitatory postsynaptic potentials (EPSPs) or inhibitory postsynaptic potentials (IPSPs). The action potentials generated or influenced by such postsynaptic potentials (PSPs) can be characterized by such metrics as frequency, temporal coincidence with events in neighboring and convergent cells, and bursting properties (action potentials generated in high frequency clusters).
Electrosensory primary afferent terminals (from A-type mormyromast receptors) and juxtalobar axonal branches (carrying the EOCD) are dense in the granular cell layer. This suggests that this layer acts as a crucial relay in the transfer of mormyromast information from afferents to Purkinje-like cells (large cell bodies, profuse dendritic branching, converts numerous inputs into a single, all-or-none spike) with projections exiting the ELL, such as the large fusiform (LF) and large ganglion (LG) cells (Meek, Grant and Bell, 1999; see figure 2). Putative granular cells that generate EPSPs (E cells) or IPSPs (I cells) have been characterized (Bell and Grant, 1992) and other preliminary data suggest that there are hetergeneous granular cell populations that either produce PSPs following the EOCD alone, or that require coincident inputs (AND gate) from both the juxtalobar EOCD and primary afferents to elicit such potentials (Meek, Grant and Bell, 1999). So, although the granular cell layer remains relatively uncharacterized, inputs from the juxtalobar nucleus, primary afferents, and LMI cells seem to be the key players (Meek et al., 1999; Bell and Grant, 1992).
To review, mormyromast afferents carry latency-coded representations of stimulus intensity from the skin, the juxtalobar input carries the EOCD, and the LMI input is thought to provide lateral inhibition and object edge sharpening in a manner similar to the horizontal and amacrine cells of the mammalian retina (figure 2). Each granular cell receives multiple afferent inputs, and each of these inputs carries information from neighboring areas of the skin (Meek et al., 1999; Bell and Grant, 1992). Because there is an array of mormyromast electroreceptors projecting to each granular cell of the ELL, and the intensity of electric field distortions depends on precise object location, each granular cell effectively has an on-center receptive field (Meek et al., 1999; Bell and Grant, 1992). Since activation of more distal receptors results in LMI inhibition of a given granular cell (figure 3), this receptive field is both on-center and off-surround (on-center in the sense that stimuli located at a preferred location will activate the corresponding granular cell, off-surround in that stimuli adjacent to this location will inhibit that cell).
{kind=link}