The Weakly Electric Fish as an Animal Model
Weakly electric fish are an ideal model for the study of sensory systems and the early processing of sensory information. These species possess three classes of sensory receptors and corresponding projection pathways specialized for the analysis of a single environmental signal, the electric field. These pathways, used for communication with conspecifics (Knollenorgan receptor), passive electrolocation (ampullary receptor), and active electrolocation (mormyromast), share the ELL as a common neural substrate. In weakly electric fish, the "visual" function of object location and navigation relies heavily on temporal information inherent in reafferent and corollary discharge signals (latency coding, gating of afferent inputs with the EOCD), where, to review, "reafferent" refers to sensory inputs originating from the fish’s own motor acts and "corollary discharge" refers to internal copies of the neural command signals initiating these motor acts (Bell, 1989; Bell and Grant, 1992) .
This makes sense from a sensory ecology viewpoint, because electric fields are relatively insensitive to temporal distortions caused by the surrounding medium (Dusenbery, 1992). Visual systems based on very high frequency electromagnetic waves, by comparison, are afforded the luxury of predominantly spatial coding because these kinds of high energy waves can maintain spatial resolution over large distances. The weak electric fields of these fish, however, have a relatively short range (figure 5) somewhere in the neighborhood of 1 to 2 meters (Heiligenberg, 1990). Weakly electric fish have evolved sensory systems that take advantage of the efficacy of using copies of their own EOD to decode the temporal characteristics of incoming sensory afferent signals, much like bats store information about the exact timing of their own vocalization and use this information to decode the time-lag of reverberations returning to their ears (Moss and Surlykke, 2001).
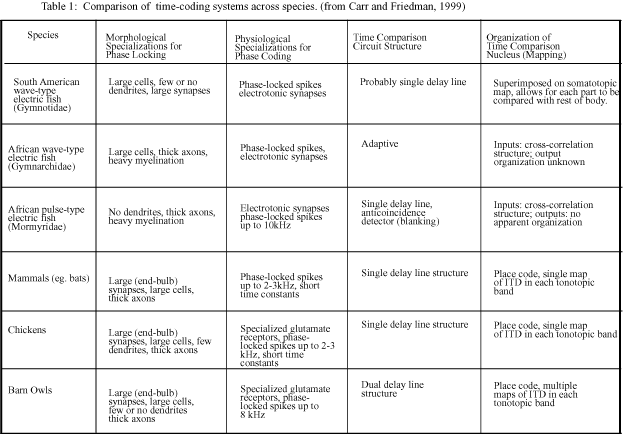
The sensory systems of the weakly electric fish provide both ready cross-modality and cross-species comparisons (Table 1). For instance, a difference between echolocation in bats and electrolocation in weakly electric fish is that the gating signal in bats is the sound of the emitted call travelling through bone tissue to the inner ear, whereas the copy in the fish is a corollary discharge signal propagated via neurons and synapses (Bell and Grant, 1992). Despite this, the function of the gating signal in both bats and weakly electric fish is to selectively enhance incoming signals that orignate from their own vocal or electric motor discharges. Therefore, isomorphism in the computations used to compute the temporal aspects of sensory inputs does not necessary imply a corresponding anatomical isomorphism.
Despite a primary interest in the workings of the human mind, we are often forced to make use of animal models because they are more accessible to invasive recordings and experimental manipulations. Therefore, it is necessary to briefly describe the utility of animal models, especially in the context of comparative neuroethology. An interest in human and primate vision, for instance, can be indirectly addressed by an approach thatcompares the neural strategies found in modalities specializing in distance perception and spatial representations across many species.
Human distance perception and object location relies on a combination of binocular and monocular cues. Disparities between retinotopic locations of an image are used to compute azimuth angle and distance of the object in relation to the viewer, and the monocular cues of contrast and blur can be used, along with other cues, to compute absolute distance from the eye (Lewis and Maler, 2001). Object distance can be computed as the ratio between contrast and blur, in that an object with high contrast and high blur could be perceived as being the same distance from the viewer as an object with low contrast and low blur, as long as the ratio between these two image qualities remained constant.
It has been shown that weakly electric fish, employing active electrolocation to navigate in the dark, rely on an analogous ratio to compute the relative and absolute distance of objects in their environments (von der Emde et al., 1998). The fish’s image of objects in its environment, coded at the skin as a 2-dimensional representation of impedences distorting its own electric field, has a maximum amplitude at the section of skin closest to the object. This peak amplitude slopes down to baseline levels near the edge of the object.
Psychophysics experiments performed on intact, behaviorally responding weakly electric fish have shown that, although their distance discrimination abilities are quite acute in normal contexts, they can be "fooled" into misjudging object distances by simple manipulation of the objects’ shapes (von der Emde et al., 1998). For instance, a spherical object at the same distance from the fish as a cube with equal volume will be perceived to be farther away (von der Emde et al., 1998). This suggests that a simple ratio of image amplitude to maximal image slope represents the minimal information that the fish requires for calculation of object distances. If we equate image amplitude and slope in the electrosensory modality with image contrast and blur in visual systems, respectively, a strong case can be made for the idea that, despite vast anatomical and morphological differences between primates and weakly electric fish, the central nervous systems of these two evolutionarily distant groups of species essentially perform the same computation to estimate distance (Lewis and Maler, 2001).
Distance estimations performed in auditory modalities work by a similar principle, with sound pressure and frequency spectrum acting as substitutes for amplitude/contrast and slope/blur. Auditory objects with dominant high frequencies and high pressure levels tend to be perceived as closer than objects with lower frequencies and lower pressures (Lewis and Maler, 2001).
Closer examination of the physiological and computational principles underlying auditory, visual, and electrosensory systems used for navigation and object location, then, is likely to yield a better understanding of the general principles of sensory systems across many species, including humans. Furthermore, consideration of the computational principles underlying these three independently evolved systems adds evolutionary insight to the problem of early coding of spatial information in the active electrosensory system of Gnathonemus petersii.
The fact that electrosensory systems have evolved independently in at least two orders of South American and African weakly electric fish, and that these systems perform similar behavioral functions with analogous but distinct neural architectures, suggests that examination of the computational algorithms underlying such behaviors, as constrained by evolution, is a justifiable approach (Carr and Friedman, 1999). Several features of sensory systems that specialize in analysis of temporal information, such as those of the barn owl and weakly electric fish, have been uncovered. These include high levels of convergence for parallel channels coding phase information, separation of channels coding phase and amplitude information, use of delay lines and topographically mapped projections, fast PSP rise-times and electrical gap junctions to preserve precise temporal information, incorporation of thick, heavily myelinated axonal projections and small, electrotonically compact somata to further preserve temporal information, and high levels of calcium-binding proteins near synaptic contacts involving glutamate receptors that mediate calcium influx (Table 1; Carr and Friedman, 1999).
A key strength of the weakly electric fish model is the practicality of in vivo and in vitro preparations and recordings, and the relative ease with which the fish’s sensory inputs may be parametrically varied. An example of this controlled, parametric stimulus control can be found in experiments exploring the jamming avoidance response (JAR) of weakly electric fish of the order Gymnotiformes (Heiligenberg, 1991). The regular, continuous nature of the Gymnotiformes’ EOD waveform makes it highly susceptible to mimicry and interference from conspecifics. Evolution has taken care of this problem with the characteristic jamming avoidance response (JAR). When a wave-emitting fish of the order Gymnotiformes encounters another fish with a nearly identical frequency waveform, it modulates its own frequency up or down to maintain the ability to distinguish its own signal. This is done by analysis of the frequency and amplitude modulations of the summated signals at different locations along the skin (Heiligenberg, 1991). Plotting frequency versus amplitude reveals a circular "beat-pulse" pattern that moves either in a clockwise or counterclockwise direction over time, depending on whether the interfering signal is of a higher or lower frequency than the fish’s own waveform (Heiligenberg, 1991).
With this phenomenon, as with most other aspects of weakly electric fish sensory perception, the electric field distortions of objects and the characteristic waveforms of the fish are easily replicated and experimentally controlled with standard electronics equipment. Also, the EOD motor command is easily blocked with a preparation of curare (nicotinic acetylcholine receptor antagonist) so that the functionality of electrosensory systems can be isolated from the effects of the electric field discharge and corresponding electrical organ corollary discharge (Bell, 1989).
Another consideration supporting the utility of the weakly electric fish as a general model for sensory processing is the predominant role that the cerebellum and cerebellar-like structures (ELL) play in these species, which have an unusually high percentage of brain mass devoted to the cerebellum and its annex, the ELL (Heiligenberg, 1990). The ELL can be viewed as an anologue to the cerebellum that is specialized for processing sensory input from the Knollenorgan, ampullary and mormyromast modalities.
Functional similarities between the neural substate for these three submodalities (the ELL) and mammalian cerebellum include the modulation of sensory inputs by expectations in the form of efference copies and corollary discharge, intrinsic feedback circuits, and feedback from higher brain areas. In a broad sense, mammalian cerebellum is a structure that uses representations of regularities in sensory input to supress redundancies and enable sparse coding in parallel fibers leading to cerebellar nuclei that feed-forward to other brain areas (Devor, 2000).
The olivary nucleus, a structure from which climbing fibers to the cerebellum originate, essentially filters out predictable sensory inputs and responds maximally to unexpected stimulation of its incoming receptive fields (Devor, 2000). This structure, in particular, appears to be the major anatomical evolutionary difference between mammalian cerebellum and the cerebellar-like structures of teleost fishes (ELL), amphibians (medial octavolateral nucleus), and mammals (for instance, the dorsal cochlear nucleus of cats integrates cross-modal inputs and appears to perform an olivary-type gating function before sending signals to higher auditory centers) (Devor, 2000). In mormyrid teleosts such Gnathonemus petersii, a function analagous to this olivary-gating is performed in the nucleus of the ELL (nELL) for Knollenorgan inputs, and in the corresponding areas of ELL cortex for both ampullary and mormyromast inputs (Bell, 1989).
The Knollenorgan system, specialized for analyzing communication between fish, is the most temporally acute of the three modalities (Bell, 1989). This acuity is maintained by morphological properties such as large, heavily myelinated axons, fast-acting electrical / gap-junction synapses, and well-defined relative delay-line lengths that control propagation speeds from one relay cell to the next (Bell, 1989). The electrical synapse eliminates delays imposed by chemical synapses, which require complex events at the membrane such as calcium-binding, vesicular fusion and release, diffusion of transmitter across the synapse, and binding to the appropriate receptors on the postsynaptic cell. In electrical synapses, current passes directly from one neuron to another through gap junctions. This direct coupling results in reduced transmission time and signal variability compared to chemically-mediated transmission. Thus, fast and reliable propagation in the Knollenorgan projection pathway, mediated by large axons, high degrees of myelination, and electrical synapses, preserves temporal properties as they are mapped from one cell layer (for instance, electroreceptors on the skin) to another cell layer (for instance, the nELL of the knollenorgan system and the granular cell layer of the mormyromast system).
The separation of channels coding for temporal and intensity information is a property of time-coding modalities across most species, including Gnathonemus petersii (Carr and Friedman, 1999). The reason for this is probably that, although both information types must ultimately be integrated further downstream for coherent stimulus representation, the morphological properties that optimize spatial and temporal acuity are for the most part mutually exclusive. For instance, increasing receptive field size and correlating a large number of receptor responses is ideal for temporal processing because jitter is minimized, but this "strategy" results in poor spatial acuity, since information about the precise origin of each input is lost through convergence (Carr and Friedman, 1999).
Since a fish’s own EOD is likely to be the strongest local electric field in its environment, due to the rapid attenuation of these static fields over short distances, communication with other fish is made possible only by filtering the reafferent signal out of Knollenorgan inputs (Bell, 1989). In Gnathonemus petersii, such filtering (the NAND gate, which performs the opposite function of the AND gate in the mormyromast system) is time-locked to EOD discharge over a narrow temporal window so that almost no inputs are lost except those evoked by the fish’s own EOD (Bell, 1989). As an instantiation of this filter, the EOCD is relatively "hard-wired" to block the corresponding reafferent input with a fast-transient IPSP occurring roughly 2ms post-EOD (Bell, 1989). The Knollenorgan blockade of reafferent inputs prevents the fish from confusing its own electrical signature with that of conspecifics (Bell et al., 1983). This is a crucial consideration, since the high spatial attenuation rate of Mormyrid electrical fields essentially guarantees that a fish’s own discharge will provide considerable interference with all fields of external origin (i.e. by drowning them out).
The ampullary system, used for passive electrolocation, shows a similar inhibition of EOD reafferent input, but the longer duration of the reafferent at lower frequencies necessitates a more plastic EOCD (Bell, 1989). The negative image, a spatial map of sensory expectations computed from recent reafferent inputs, is subtracted from the total ampullary input, and this negative image is updated with each EOD (Bell, 1989). While the ampullary system is useful for electrolocation based on the electric fields of external origin, it cannot provide information about objects that do not create electric fields. For this the fish relies on active electrolocation mediated by mormyromast electroreceptors and the corresponding ELL projection pathway, which we will now consider in the context of a computer model.
{kind=link}
{kind=link}