Granular Cell Integration and Conversion of Latency-to-Fire Code into a Burst Duration Code: Modeling in the NEURON Simulation Environment
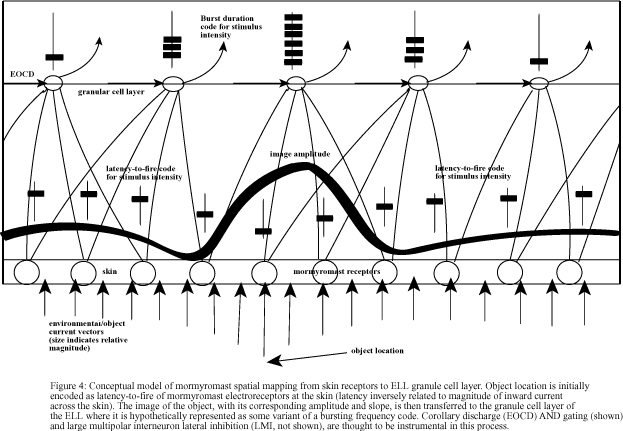
The current mathematical model is intended to explore the ionic conductances, input connectivities and postsynaptic events that could mediate a transformation of the latency code for stimulus intensity in primary mormyromast afferents into a burst duration code in granular cells of the medial ELL. Primary afferent EPSPs arriving with post-EOCD delays brief enough to trigger granular cell spikes are expected, based on the preliminary electrophysiological recordings, to produce a graded response encoded as the number of spikes in the granular cell burst following the arrival of each successive EOCD (Bell and Grant, 1992; also, figure 4). It is expected that decreases in afferent latency, signaling increases in stimulus intensity, will produce corresponding increases in the duration of granular cell bursts.
Knowledge of the brain evolves through exploitation of the similarities that exist across various species and structures that employ heterogeneous anatomical structures to perform similar computational tasks involved in "making sense" of sensory inputs. With this in mind, examination of recent models of cerebellar granule cell bursting and resonance properties (D’Angelo et al., 2001), and cerebellar granule cell coincidence detection in the turtle (Gabbiani et al., 1994), will aid in conceptualization of the present model addressing the transformation of a latency-to-fire code for stimulus intensity in mormyromast primary afferent fibers into a burst duration code in granular cells of the medial ELL in the weakly electric fish, Gnathonemus petersii.
Subthreshold voltage fluctuations mediated by various potassium (K+) and sodium (Na+) currents across the neuronal cell membrane exert significant influence on the bursting properties of the neuron (D’Angelo et al., 2001). These fluctuations in granule cells tend to be regular and periodic, with the resting membrane potential oscillating about a fixed point (typically —65 to —80mV) at frequencies of 3-12 Hz. Resonance in this range, termed theta-frequency, is driven primarily by a slow repolarizing K+ current (I-KM) in combination with a persistent Na+ current (I-NaP). The hyperpolarizing effect of this K+ current, combined with the prolonged depolarizing force exerted by I-NaP, produces a dynamic membrane voltage equilibrium both at subthreshold voltage ranges and during bursts of action potentials.
The phasic subthreshold fluctuations in membrane potential that result from the interaction of these two currents, as described by D’Angelo et al. (2001), may play an active role in controling the temporal gating function that the EOCD performs on mormyromast afferent inputs arriving at the granular cell layer in the present model. However, these conductance dynamics and intuitions about subthreshold membrane voltage activity need to be complemented with consideration of realistic synaptic inputs to the model cell.
A model of coincidence detection in cerebellar granule cells of the turtle (Gabbiani, Midtgaard and Knopfel, 1994) provides predictions concerning the network properties that are a necessary extension of these single-cell dynamics. The key similarities between the cerebellar granule cells of the turtle and the ELL granular cells of the mormyromast modality of Gnathonemus petersii, as they are currently understood, include degree of input convergence (4-5 mossy fiber inputs to each turtle granule cell, 3-4 afferent inputs to each ELL granular cell), conceptual function (selective filtering to produce a more robust representation of a particular stimulus in the form of a bursting frequency code), conductances and receptors present at the synapses and somata of both cell types (which include a fast sodium conductance, voltage-gated calcium conductance, delayed potassium conductance, mixed NMDA/AMPA synapses, etc…), and shunting inhibition (lateral inhibition provided by Golgi cells in the turtle model, LMI cells in the current model) at the soma (Gabbiani, Midtgaard and Knopfel, 1994).
Results of the Gabbiani et al. (1994) model of turtle cerebellar granule cell integration suggest that temporal precision in the decoding and filtering of inputs to the granule cell depends heavily on the modulation of NMDA receptor-mediated EPSPs by the shunting, GABAergic inhibitory inputs (Gabbiani et al., 1994). A combination of AMPA-mediated EPSPs, which have extremely fast rise and decay time constants, NMDA-mediated EPSPs, which are by comparison much slower to decay, and shunting inhibition that effectively blocks some portion of the tail of the NMDA-mediated EPSPs, is both predicted by the turtle model and consistent with observations of the ELL granular cell layer (Gabbiani et al., 1994; Bell and Grant, 1992). It is therefore reasonable that such a scheme is implemented in both systems.
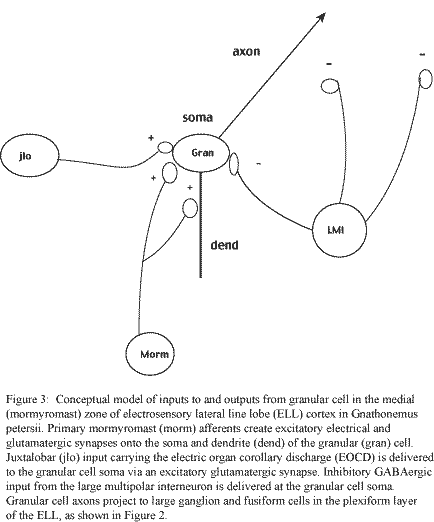
Due to the difficulties in recording from ELL granular cells, which have somata merely 5 micrometers in diameter, relevant electrophysiological inquiries have so far been limited to extracellular field recordings in the granular cell layer and intracellular recordings of LMI, juxtalobar (EOCD), and primary mormyromast afferents. These data, combined with knowledge of morphology and connectivity taken from stainings of calbindin (Meek, unpublished observations), a calcium-binding protein ubiquitous in these cells, and the more macroscopic characterization of behaviors involving active electrolocation, provide the empirical basis on which the model will be tested (figure 3).
The model is a second-order conceptual tool, allowing for the development of an internally consistent theory accounting for the biological mechanisms underlying active electrolocation in the Mormyrid weakly electric fish, Gnathonemus petersii. Although initial results of the model are sensitive to deviations due to somewhat arbitrary parameter specifications, coevolution with the experimental investigations it suggests will ensure adequate biological realism. Under the scheme drawn by Abbot and Marder (1995), this coevolution will entail confirmatory functions, by testing whether extant morphological and electrophysiological data are consistent with the integrative conceptual understanding provided by the model, and speculative functions, by making predictions that are testable both in vivo and in vitro. The model may also be viewed as a sophisticated form of data analysis, an interpretive engine facilitating bidirectional interactions between quantitative theory and empirical data.
{kind=link}
{kind=link}