Chapter III: Results
Overview
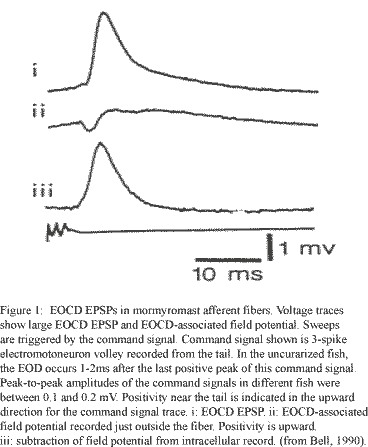
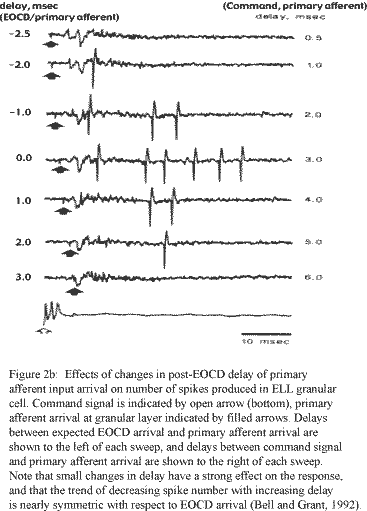
Although experimental data providing constraints on the model were limited in many respects, several key restrictions were applied. These included the requirement of both electrical and glutamatergic inputs from the primary afferent onto the granular cell, glutamatergic input from the juxtalobar fiber onto the granular cell, a lower limit on the decay time and amplitude of the EPSP produced by the EOCD juxtalobar input to the granular cell (figure 1), and a temporally strict dependence of spike number output on the relative timing of inputs from these two cells (figures 2a, 2b). Other elements of the model, however, were allowed to vary to produce results consistent with the theoretical coding transformation of latency into spike number.
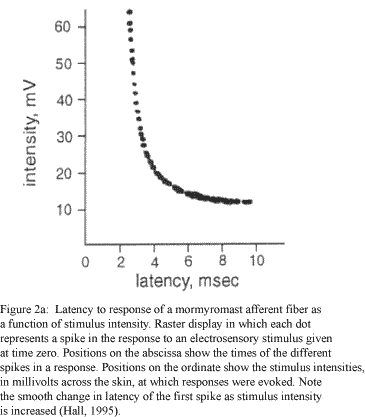
The physiological dynamic range of post-stimulus firing latencies in the primary afferent fiber is around 6ms, where a latency of 0ms would produce a granular cell input directly coincident with the arrival of the juxtalobar EOCD (figure 2b, Bell and Grant, 1992). Additionally, the number of spikes generated in the granular cell following coincident arrival of the juxtalobar EOCD and primary afferent inputs was six when the amplitude of the primary afferent PSP was set to produce a spike number as function of delay curve that decreased to zero at +/- 3ms with respect to the arrival of the EOCD input (figure 2b, Bell and Grant, 1992). Although these results from extracellular field recordings do not represent the complete range of responses for all granular cells of the medial ELL (some cells showed responses following the arrival of primary afferent impulses up to 30ms post-EOCD), the maximum offset latency (~50ms) and spike number of the granular cell response under these conditions (Bell and Grant, 1992) were used as initial constraints on the model.
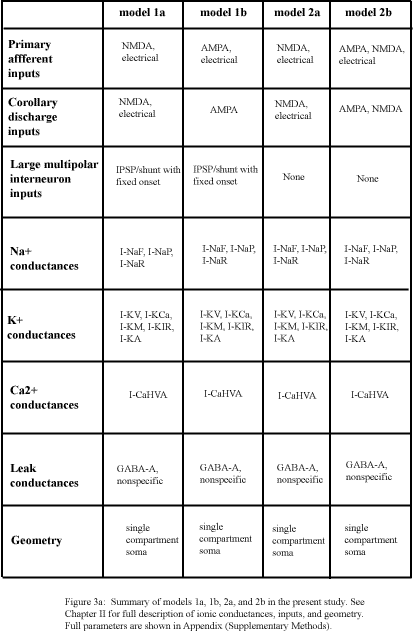
Following initial testing of a single compartment model incorporating default ionic conductances from the D’Angelo et al (2001) model of rat cerebellar granular cells referred to as the preliminary model, two major classes of models were created. The first class of model (models 1a and 1b; figure 3) was created to maximize the fit to a theoretical latency to spike number transformation for a single compartment granular cell. This model, despite achieving a nearly linear inverse relationship between input delay and spike number, required use of input characteristics that have yet to be identified in vivo. The second class of model (models 2a and 2b; figure 3) was built to retain the fit to theoretical coding constraints while gradually eliminating speculative input characteristics. In this case, the desired relationship between spike number and input delay was achieved only by allowing the time window of the total granular cell response to extend over a time window of approximately 4-fold greater duration than has been observed in vivo.
The input PSPs and maximum ionic conductances (figure 3) of both models were systematically varied to explore the parameter space suggested by expermental data. In addition, sensitivities to manipulation of maximum ionic conductances for the included ion channels were determined and are reported here. Total voltage and current responses of simulated granular cells are also presented to provide an integrative understanding of the mechanism of latency to spike number transformation.
{kind=link}
{kind=link}
{kind=link}
{kind=link}